Да би се свеобухватно могло говорити о хемоглобину (Хб), корисно је прво се позабавити миоглобином (Мб) који је врло сличан хемоглобину, али је много једноставнији. Постоје строги сроднички односи између хемоглобина и миоглобина: оба су коњугована протеина и њихова протетска група (не-протеински део) је хеме група.
Миоглобин је глобуларни протеин који се састоји од једног ланца од око стотину и педесет аминокиселина (у зависности од организма) и његова молекуларна тежина је око 18 Кд.
Као што је поменуто, он је опремљен хеме групом која је убачена у хидрофобни (или липофилни) део протеина, који се састоји од набора који се могу приписати α-хеликс структури фиброзних протеина.
Миоглобин се састоји углавном од сегмената α-хеликса, присутних у броју осам и састоји се, готово искључиво, од неполарних остатака (леуцин, валин, метионин и фенилаланин) док су поларни остаци практично одсутни (аспарагинска киселина, глутаминска киселина, лизин и аргинин); једини поларни остаци су два хистидина, који играју фундаменталну улогу у нападу кисеоника на хеме групу.
Хеме група је хромофорска група (апсорбује се у видљивом) и представља функционалну групу миоглобина.
Види такође: гликирани хемоглобин - хемоглобин у урину
Мало хемије
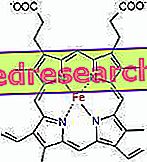
Веза између протопорфирина и гвожђа је типична веза такозваних координационих једињења која су хемијска једињења у којима централни атом (или ион) формира везе са другим хемијским врстама у броју који је већи од његовог оксидационог броја (електрични набој). У случају хема, ове везе су реверзибилне и слабе.
Координациони број (број координационих веза) гвожђа је шест: око гвожђа може бити шест молекула који деле електроне који се вежу.
Да би се формирало координационо једињење, потребно је две орбитале са правилном оријентацијом: једна која може да "купи" електроне, а друга да их донира
У хему, гвожђе формира четири планарне везе са четири атома азота који су у центру прото-порфиринског прстена и петом везом са проксималним хистидинским азотом; гвожђе има шесту слободну координациону везу и може се везати за кисик.
Када је гвожђе у облику слободног иона, његове орбитале типа д имају исту енергију; у миоглобину, јон гвожђа је везан за протопорфирин и хистидин: ове врсте магнетски ремете орбитале гвожђа; обим поремећаја ће бити различит за различите орбитале у складу са њиховом просторном оријентацијом и оном ометајућих врста. Пошто се укупна енергија орбитала мора вратити на константу, пертурбација узрокује енергетско раздвајање између различитих орбитала: енергија добијена неким орбиталима је еквивалентна енергији коју су изгубили други.
Ако раздвајање које се дешава између орбитала није јако велико, пожељан је електронски аранжман са високим спин-ом: електрони за везивање покушавају да се распореде у паралелним спиновима на неколико могућих под-нивоа (максимална мултиплицитет); ако је, с друге стране, пертурбација веома јака и постоји велика раздвојеност између орбитала, може бити прикладније упарити електроне везивања у орбитале ниже енергије (ниски спин).
Када се гвожђе везује за кисеоник, молекул преузима ниску диспензију спина, док када гвожђе има шесто слободно координационо повезивање, молекул има високи спин.
Захваљујући тој разлици у спину, кроз спектралну анализу миоглобина, можемо да разумемо да ли постоји везани кисеоник (МбО2) или не (Мб).
Миоглобин је протеин карактеристичан за мишиће (али не само у мишићима).
Миоглобин је екстрахован из кита у којем је присутан у великим количинама и затим пречишћен.
Китови имају дисање као људска бића: имају плућа да апсорбују ваздух кроз респираторни процес; кит-сперматозоид мора носити што је више могуће кисика у мишићима који су способни да акумулирају кисеоник тако што га вежу за присутни миоглобин; Кисеоник се затим полако ослобађа када се китови урони због тога што његов метаболизам захтева кисеоник: што је већа количина кисеоника коју кит може да апсорбује и више кисеоника је доступно током роњења.

Да би месо било мање или више црвено је садржај хемопротеина (то је хеме које чини месо црвеном).
Хемоглобин има многе структурне аналогије са миоглобином и способан је да реверзибилно веже молекуларни кисеоник; али, иако је миоглобин ограничен на мишиће и периферна ткива уопште, хемоглобин се налази у еритроцитима или црвеним крвним зрнцима (то су псеудо-ћелије, тј. оне нису праве ћелије) које чине 40% крви.
Хемоглобин је тетраметар, тј. Састоји се од четири полипептидна ланца, сваки са хеме групом и два по два (у људском бићу постоје два алфа ланца и два бета ланца).
Главна функција хемоглобина је транспорт кисеоника ; Друга функција крви у којој је укључен хемоглобин је транспорт супстанци до ткива.
На путу од плућа (богат кисеоником) до ткива, хемоглобин преноси кисеоник (у исто време и друге супстанце такође допиру до ткива) док на обрнутом путу носи са собом отпад сакупљен из ткива, посебно угљен диоксид који настаје у метаболизму.
У развоју људског бића постоје гени који се изражавају само одређени временски период; из тог разлога имамо различите хемоглобине: фетални, ембрионски, одраслог човека.
Ланци који сачињавају ове различите хемоглобине имају различите структуре, али са сличностима функција коју обављају више или мање је иста.
Објашњење присуства неколико различитих ланаца је следеће: током еволутивног процеса организама, хемоглобин се такође развио специјализован за транспорт кисеоника из подручја која су богата у дефицитарним подручјима. На почетку еволуционог ланца хемоглобин је преносио кисеоник у малим организмима; у току еволуције организми су достигли веће димензије, па се хемоглобин променио да би могао да транспортује кисеоник у подручја даље од тачке да је богат; да би се то урадило, нове структуре ланаца које чине хемоглобин су кодификоване током еволуционог процеса.
Миоглобин везује кисеоник чак и при скромним притисцима; у периферним ткивима постоји притисак (ПО2) од око 30 ммХг: на овом притиску миоглобин не ослобађа кисеоник, тако да би био неефикасан као носач кисеоника. Хемоглобин, с друге стране, има више еластично понашање: веже кисеоник при високим притисцима и ослобађа га када притисак опада.
Када је протеин функционално активан, може благо да промени свој облик; на пример, оксигенисани миоглобин има другачији облик од не-оксигенисаног миоглобина и ова мутација не утиче на суседне.
Дискурс је другачији у случају повезаних протеина као што је хемоглобин: када је ланац оксигенисан, индукује се да промени свој облик, али ова модификација је тродимензионална, тако да су и други ланци тетраметра такође погођени. Чињеница да су ланци повезани једни с другима доводи до тога да мисли да модификација једног утиче на друге сусједне, чак и ако је у различитој мјери; када је ланац оксигенисан, други ланци тетраметра имају "мање непријатељски став" према кисеонику: потешкоћа са којом ланчани оксигенати падају како се ланци поред њега оксидирају сами. Исти аргумент важи и за деоксигенацију.
Квартарна структура деоксихемоглобина назива се Т (напета) форма, док се оксихемоглобин назива Р облик (ослобођен); у растегнутом стању постоји низ прилично јаких електростатских интеракција између киселинских аминокиселина и основних аминокиселина које доводе до ригидне структуре деоксихемоглобина (због тога је "напета форма"), док је када је кисеоник везан, ентитет интеракције се смањују (због тога је "ослобођена форма"). Даље, у одсуству кисеоника, хистидински набој (види структуру) стабилизован је супротним набојем аспарагинске киселине, док у присуству кисеоника постоји тенденција да протеин губи протон; све то значи да је оксигенирани хемоглобин јача киселина од деоксигениране хемоглобије: бохров ефекат .
У зависности од пХ вредности, хеме група се мање или више лако везује за кисеоник: у киселој средини хемоглобин лакше ослобађа кисеоник (напета форма је стабилна), док је у базичној средини веза са кисеоником јачи.

Сваки хемоглобин ослобађа 0.7 протона по долазном молу кисеоника (О2).
Бохров ефекат омогућава хемоглобину да побољша своју способност да носи кисеоник.
Хемоглобин који путује од плућа до ткива мора се уравнотежити у складу са притиском, пХ и температуром.
Видимо ефекат температуре .
Температура у пулмонарним алвеолама је око 1-1, 5 ° Ц нижа од спољне температуре, док је у мишићима температура око 36, 5-37 ° Ц; Како се температура повећава, фактор засићења се смањује (са истим притиском): то се дешава зато што се кинетичка енергија повећава и дисоцијација је фаворизована.

Постоје и други фактори који могу утицати на способност хемоглобина да се веже за кисеоник, од којих је једна концентрација 2, 3-бисфосфоглицерата .
2, 3-бисфосфоглицерат је метаболички агенс присутан у еритроцитима у концентрацији од 4-5 мМ (ни у једном другом делу тела није присутан у тако високој концентрацији).
При физиолошком пХ, 2, 3-бисфосфоглицерат се депротонира и на њему има пет негативних набоја; она иде у клин између два бета ланца хемоглобина, јер ови ланци имају високу концентрацију позитивних набоја. Електростатичке интеракције између бета ланаца и бисфосфоглицератних ланаца дају одређену крутост систему: добива се напета структура која има мали афинитет за кисеоник; током оксигенације, онда је 2, 3 бисфосфоглицерат избачен.
У еритроцитима постоји специјални апарат који 1, 3-бисфосфоглицерат (произведен метаболизмом) претвара у 2, 3-бисфосфоглицерат тако да достигне концентрацију од 4-5 мМ и због тога хемоглобин може да замени кисеоник у ткивима.
Хемоглобин који стиже у ткиво је у ослобођеном стању (везан за кисеоник), али у близини ткива је карбоксилисан и прелази у растегнуто стање: протеин у овом стању има мању тенденцију да се веже са кисеоником, у поређењу са ослобођеним стањем, хемоглобин ослобађа кисеоник у ткиво; Надаље, због реакције између воде и угљичног диоксида долази до производње Х + иона, дакле даљњег кисика због ефекта бохра.
Угљен диоксид дифундира у еритроцит који пролази кроз плазма мембрану; с обзиром да еритроцити чине око 40% крви, требало би очекивати да само 40% угљендиоксида који дифундује из ткива улази у њих, у стварности 90% угљичног диоксида улази у црвене ћелије јер садрже ензим који се претвара угљен диоксид у угљеној киселини, следи да је стационарна концентрација угљен-диоксида у еритроцитима ниска и да је стога брзина уласка висока.

Још једна појава која се јавља када еритроцит дође до ткива је сљедећи: градијентом, ХЦО3- (дериват угљичног диоксида) излази из еритроцита и, да би уравнотежио излаз негативног набоја, имамо унос хлорида који одређује повећање осмотског притиска: да би се балансирала ова варијација долази и до уласка воде која узрокује отицање еритроцита (ефекат ХАМБУРГЕР). Супротни феномен се јавља када еритроцит досегне плућне алвеоле: долази до дефлације еритроцита (ефекат ХАЛДАНЕ). Тако су венски еритроцити (директно у плућа) округлији од артеријских.


